Последние материалы
- Уровень насилия на Ближнем Востоке в бронзовом веке был ниже, чем в медном и железном
- Нобелевская премия по физике — 2023
- Литиевое месторождение в древней кальдере вулкана Йеллоустон образовалось в результате повторной магматической активизации
- Неандертальцы охотились на пещерных львов и использовали их шкуры
- Планарии несколько раз приобретали и теряли способность к регенерации головы
- Новая реконструкция девонской рыбы дунклостея уменьшила его длину и увеличила массу
- Летучие мыши освоили эхолокацию не меньше 50 миллионов лет назад
- Пересадка крысам кишечной микрофлоры больных Альцгеймером вызвала у них симптомы этой болезни
- Импактная гипотеза формирования Луны объясняет неоднородность земной мантии
- Нобелевская премия по химии — 2023
| Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было |
| 23.11.2017 13:47 |
 Пятьдесят лет назад, в 1967 году, Линн Маргулис (Lynn Margulis) опубликовала развернутое изложение симбиогенетической теории, согласно которой эукариоты (организмы с клеточными ядрами) возникли в результате серии объединений разных клеток между собой. Современная поправка к этой теории гласит, что в основе становления эукариот, по-видимому, была не общая тенденция, охватившая многие эволюционные ветви (как предполагала Маргулис), а уникальное событие, приведшее к слиянию клеток археи и протеобактерии. В результате образовалась сложная клетка с митохондриями, которая и стала первым эукариотом. Дальнейшие симбиогенетические события — например, захват водорослей, ставших хлоропластами, — действительно происходили много раз, но с возникновением эукариот как таковых они не связаны. Пятьдесят с лишним лет назад, в марте 1967 года, в международном «Журнале теоретической биологии» (Journal of Theoretical Biology) вышла статья «О происхождении клеток, делящихся митозом» (L. Sagan, 1967. On the origin of mitosing cells). Автора статьи звали Линн Саган (Lynn Sagan), но в дальнейшем эта замечательная женщина стала гораздо более известна как Линн Маргулис (Lynn Margulis). Фамилию Саган она носила, потому что была некоторое время замужем за Карлом Саганом (Carl Edward Sagan), астрономом и писателем. Выход в 1967 году статьи Линн Маргулис (будем для удобства звать ее так) стал началом обновления биологических представлений, которое многие авторы расценили как смену парадигм — то есть, иными словами, как самую настоящую научную революцию (И. М. Мирабдуллаев, 1991. Эндосимбиотическая теория — от фантастики к парадигме). Суть интриги тут проста. Со времен Чарльза Дарвина биологи были убеждены, что основным способом эволюции является дивергенция — расхождение ветвей. Линн Маргулис была первой, кто сумел по-настоящему убедительно объяснить научному сообществу, что механизм некоторых крупных эволюционных событий, скорее всего, был принципиально другим. В центре интересов Маргулис оказалась проблема происхождения эукариот — организмов, клетка которых обладает сложной внутренней структурой с ядром. К эукариотам относятся животные, растения, грибы и многие одноклеточные — амебы, жгутиконосцы, инфузории и прочие. Маргулис показала, что ранняя эволюция эукариот вовсе не сводилась к дивергенции — она включала в себя слияние эволюционных ветвей, причем неоднократное. Дело в том, что по меньшей мере два типа эукариотных органелл — митохондрии, благодаря которым мы можем дышать кислородом, и хлоропласты, осуществляющие фотосинтез, — происходят не от того предка, что основная часть эукариотной клетки (рис. 1). И митохондрии, и хлоропласты — это бывшие бактерии, изначально совсем не родственные эукариотам (протеобактерии в случае митохондрий и цианобактерии — в случае хлоропластов). Эти бактерии были поглощены клеткой древнего эукариота (либо предка эукариот) и продолжили жить внутри нее, сохраняя до поры собственный генетический аппарат. Таким образом, эукариотная клетка — это, по выражению Маргулис, мультигеномная система. И возникла она в результате симбиоза, то есть взаимовыгодного сожительства разных организмов (точнее, эндосимбиоза, один из участников которого живет внутри другого). Соответствующие эволюционные ветви при этом, разумеется, слились. Такой взгляд на эволюцию получил название теории симбиогенеза. Сейчас теория симбиогенеза общепринята. Она подтверждена настолько строго, насколько вообще можно подтвердить какую бы то ни было теорию, касающуюся крупномасштабной эволюции. Но научные концепции, в отличие от религиозных догматов, никогда не остаются статичными. Естественно, что общая картина симбиогенеза выглядит для нас сейчас не совсем так (а местами и совсем не так), как представляла ее себе Линн Маргулис полвека назад. Логика классикаК пятидесятилетию выхода знаменитой статьи о симбиогенезе Journal of Theoretical Biology подготовил специальный выпуск, целиком посвященный творческому наследию Линн Маргулис. В этот выпуск входит обстоятельная статья известного британского биохимика и популяризатора науки Ника Лейна (Nick Lane), в которой современное состояние проблемы происхождения эукариот сравнивается с классическими идеями на эту тему. Лейн нисколько не сомневается, что в основных утверждениях (касающихся происхождения митохондрий и хлоропластов) Маргулис была права; в наше время в этом не сомневается, кажется, никто из серьезных ученых, ибо данные молекулярной биологии на этот счет однозначны. Но дьявол, как известно, живет в деталях. В данном случае мы можем, погрузившись в детали, найти там много нового и интересного, а главное — убедиться, что тема происхождения эукариот далеко не исчерпана. Начнем с того, что кое-какие частные предположения Маргулис оказались неверны. Это нормально: учитывая огромную скорость развития биологии, просто невероятно, чтобы в статье, опубликованной полвека назад, было точно угадано абсолютно всё. Новые факты, которые не могли быть в свое время известны автору, обязательно внесут какие-нибудь коррективы. Так получилось и тут. Прежде всего, Маргулис настаивала на симбиотическом происхождении не только митохондрий и хлоропластов, но и эукариотных жгутиков. Она считала, что предками жгутиков были закрепившиеся на эукариотной клетке длинные спирально закрученные подвижные бактерии, похожие на современных спирохет (см. рис. 1). Увы, эта гипотеза не получила никаких молекулярно-биологических подтверждений, и сейчас ее больше никто не поддерживает. В некоторых моментах Маргулис могла бы оказаться права (это не запрещено ни законами природы, ни внутренней логикой ее собственной теории), но тем не менее по не зависящим от нее причинам промахнулась. Например, она считала, что раз уж митохондрии — потомки бактерий, то рано или поздно биологи научатся культивировать их в питательной среде вне эукариотных клеток — ну, как обычных микробов. Если бы такое оказалось возможным, это было бы идеальным доказательством теории симбиогенеза. Увы, на самом деле современные митохондрии принципиально неспособны к самостоятельному выживанию, потому что большая часть их генов в ходе эволюции мигрировала в клеточное ядро и встроилась там в геном эукариотного «хозяина». Теперь белковые продукты этих генов синтезируются за пределами митохондрии, а потом переправляются в нее с помощью особых транспортных систем, принадлежащих эукариотной клетке. Гены, оставшиеся в самой митохондрии, всегда малочисленны — для жизнеобеспечения их не хватит. В 1967 году этого просто еще никто не знал. Однако по большому счету всё это частности. Мышление Линн Маргулис было синтетическим: она не ограничивалась объяснениями отдельных фактов, а стремилась свести их в целостную систему, описывающую эволюцию живых организмов в контексте истории Земли (рис. 2). Современные научные знания позволяют проверить эту систему представлений на прочность.  Рис. 2. Графическая схема эволюции жизни на Земле, примерно соответствующая взглядам Линн Маргулис. К двум центральным факторам — естественному отбору и симбиогенезу — тут добавлено действие сил, берущих начало от внутреннего тепла Земли («динамика Земли»). Протистами в принятой здесь терминологии называются все эукариоты, не относящиеся ни к животным, ни к растениям, ни к грибам. LUCA — последний общий предок всех живых клеток (Last Universal Common Ancestor). Разумеется, это древо не стоит воспринимать как истину в последней инстанции: местами оно слишком упрощено (например, проигнорировано различие между бактериями и археями), а некоторые датировки событий можно оспорить, благо палеонтология их постоянно корректирует; но в качестве более или менее адекватной общей картины оно годится. Обратим внимание, что симбиогенез тут выделен как целый принципиально важный эволюционный этап. Иллюстрация из статьи U. Kutschera, 2013. From the scala naturae to the symbiogenetic and dynamic tree of life, с изменениями Древо и сетьВсё началось с кислорода. В древнейшей атмосфере Земли молекулярного кислорода (O2) не было. Потом цианобактерии, первыми освоившие кислородный фотосинтез, стали выделять этот газ в атмосферу (для них он был просто ненужным побочным продуктом). Между тем чистый кислород — это весьма ядовитое вещество для всех, у кого нет специальных биохимических средств защиты от него. Неудивительно, что выбросы кислорода цианобактериями отравили атмосферу Земли и привели к массовому вымиранию. Начался «кислородный холокост» (L. Margulis, D. Sagan, 1997. Microcosmos: four billion years of microbial evolution). Тут уже необходима поправка. Многие современные исследователи считают, что переход от бескислородной биосферы к кислородной на самом деле был гораздо более постепенным и менее разрушительным, чем предполагают рассуждения о «кислородном холокосте» (см., например: «Великое кислородное событие» на рубеже архея и протерозоя не было ни великим, ни событием, «Элементы», 02.03.2014). Более того, не исключается, что появление свободного кислорода скорее даже повысило разнообразие микроорганизмов, потому что окисление атмосферным кислородом ряда минералов обогатило химический состав среды и создало новые экологические ниши (M. Mentel, W. Martin, 2008. Energy metabolism among eukaryotic anaerobes in light of Proterozoic ocean chemistry). В общем, представление о появлении кислорода в атмосфере как о разовой грандиозной катастрофе, поделившей всю историю Земли на «до» и «после», теперь, похоже, устарело. Так или иначе, несомненно, что больше всех от обогащения нашей планеты кислородом выиграли альфа-протеобактерии. Они научились непосредственно использовать кислород для получения энергии — причем с огромной эффективностью. А вот у одноклеточных предков эукариот такой способности не было. Они были анаэробными, то есть дышать кислородом не умели. Зато они были хищниками, научившимися поглощать более мелкие клетки путем фагоцитоза. И это дало им превосходную возможность: захватывать некоторых бактерий, не переваривая их, а «порабощая» и присваивая продукты их обмена веществ. Поглотив альфа-протеобактерию, примитивный эукариот получил возможность дышать кислородом — так образовались митохондрии. А поглотив цианобактерию, он получил возможность фотосинтезировать — так образовались хлоропласты. Маргулис считала, что такие события происходили много раз, подчиняясь возникшей общей тенденции. Это — так называемый сценарий сериального эндосимбиоза. Итак, у Маргулис получается, что на определенном этапе развития жизни эндосимбиоз стал едва ли не всеобщей закономерностью. Тогда в основании эволюционного древа эукариот должна находиться буквально целая сеть эволюционных ветвей, пересекающихся друг с другом за счет эндосимбиотических событий и «растущих» примерно в одном направлении — в том, которое диктовалось сочетанием тогдашних внешних условий со структурными особенностями клеток (рис. 3, А). 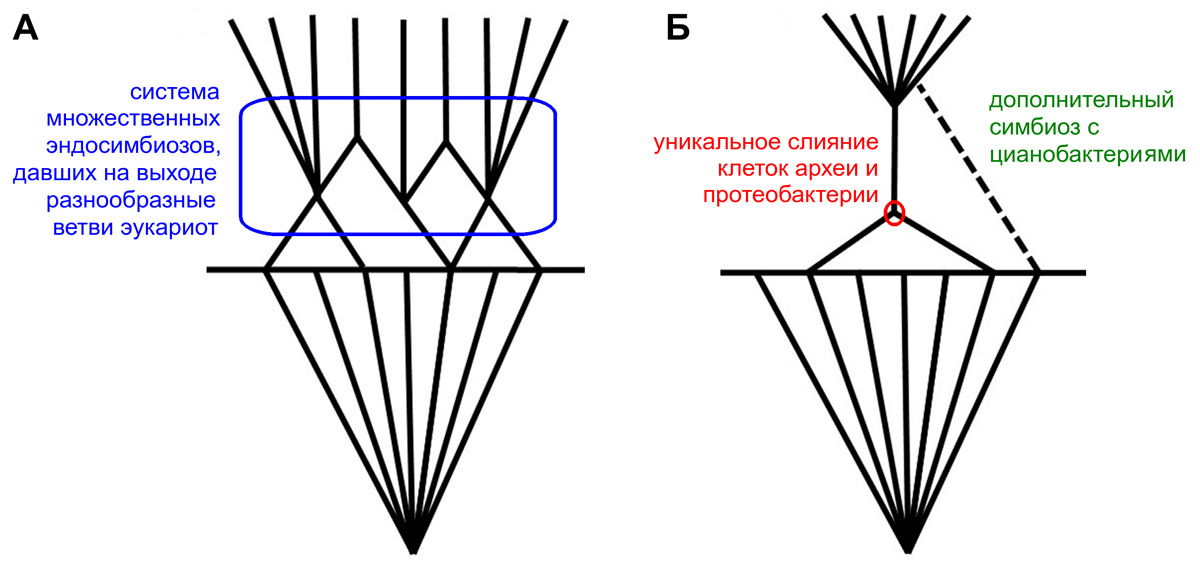 Рис. 3. Сценарии эукариотизации по Маргулис (А) и по Лейну (Б): параллельная эукариотизация versus «бутылочное горлышко». Пояснения в тексте. Иллюстрация из обсуждаемой статьи в Journal of Theoretical Biology, с изменениями Надо сказать, что к концу XX века в эволюционной биологии (и особенно в палеонтологии) и без того завоевала определенную популярность идея, что большинство крупных эволюционных событий имеет закономерный и системный характер. Подобное событие охватывает сразу много эволюционных ветвей, в которых под действием общей наследственности параллельно возникают примерно одни и те же признаки (см., например: А. Г. Пономаренко, 2004. Артроподизация и ее экологические последствия). Примерами таких событий называли маммализацию (происхождение млекопитающих), ангиоспермизацию (происхождение цветковых растений), артроподизацию (происхождение членистоногих), тетраподизацию (происхождение наземных позвоночных), орнитизацию (происхождение птиц) и многое другое. Казалось, что становление эукариот — эукариотизация — великолепно вписывается в этот ряд. Например, Кирилл Еськов в своей замечательной книге «История Земли и жизни на ней» (написанной в 1990-е годы) говорит следующее: «Скорее всего, различные варианты эукариотности, то есть внутриклеточных колоний, возникали многократно (например, есть основания полагать, что красные водоросли, резко отличающиеся от всех прочих растений по множеству ключевых признаков, являются результатом такой “независимой эукариотизации” цианобактерий)» (К. Ю. Еськов, 2000. История Земли и жизни на ней). Увы, применительно к эукариотам (прочие примеры «-заций» мы сейчас не обсуждаем) современные данные ставят этот красивый сценарий под сомнение. Проблема митохондрийНачнем с того, что обсуждавшаяся Еськовым гипотеза насчет красных водорослей теперь устарела. Молекулярные исследования показывают, что эволюционная линия красных водорослей находится глубоко внутри древа эукариот (они достаточно близкие родственники зеленых растений), и их независимая эукариотизация крайне маловероятна. Но гораздо серьезнее другое. Если симбиогенез был закономерным, долгим, многоступенчатым процессом, да еще и шел параллельно в разных эволюционных ветвях, то следовало бы ожидать, что мы увидим спектр довольно разнообразных переходных состояний между эукариотами и не-эукариотами. Маргулис именно так и думала. То, что эти переходные состояния не бросаются в глаза, она (насколько можно судить) считала проблемой чисто технической, связанной с недостатком знаний и несовершенством методов. Подтверждается ли это сейчас, когда мы знаем о живых клетках неизмеримо больше, чем знали пятьдесят лет назад? Порассуждаем. Предполагаемый сериальный эндосимбиоз должен был идти, во-первых, постепенно, и во-вторых — немного по-разному в разных эволюционных линиях (поскольку точных повторений в эволюции не бывает). Исходя из этого, Маргулис предсказывала, что рано или поздно будут обнаружены эукариоты, имеющие хлоропласты, но никогда не имевшие митохондрий; эукариоты, сохранившие бактериальные жгутики (которые резко отличаются по структуре от жгутиков эукариот); и наконец, первично анаэробные эукариоты, в клетках которых нет никаких следов приспособления к кислородной атмосфере. Ни одно из этих предсказаний не подтвердилось. Ни у кого из эукариот нет и намека на жгутики бактериального типа — средства движения у них совсем другие. Никого из известных эукариот нельзя назвать первичным анаэробом — все они, без исключения, прошли когда-то в своей эволюции «кислородную фазу». Наконец, у всех эукариот есть или действующие митохондрии, или их остатки, потерявшие значительную часть функций (гидрогеносомы, митосомы), или — на худой конец — митохондриальные гены, успевшие перейти в ядро. В конце XX века была популярна гипотеза, что у некоторых современных одноклеточных эукариот митохондрий нет и не было никогда. Таких первично безмитохондриальных эукариот предлагали выделить в особое царство Archezoa. Маргулис довольно рано приняла эту гипотезу и была верна ей до последнего — даже тогда, когда ее уже отвергли многие другие ученые (L. Margulis et al., 2005. “Imperfections and oddities” in the origin of the nucleus). Она считала вполне вероятным, что первично безмитохондриальные эукариоты («архепротисты») до сих пор живут в каких-нибудь труднодоступных бескислородных местообитаниях, где их очень сложно обнаружить. Увы, никаких «архепротистов» до сих пор так и не нашли, а вот остатков митохондрий у тех одноклеточных, которых раньше относили к Archezoa, найдено сколько угодно. На данный момент известен только один эукариот, не имеющий вообще никаких следов митохондрий, — жгутиконосец Monocercomonoides, но положение этого существа на эволюционном древе не оставляет сомнений в том, что и у него митохондрии когда-то были (A. Karnkowska et al., 2016. A eukaryote without a mitochondrial organelle). В общем, на данный момент все без исключения случаи отсутствия митохондрий у эукариот приходится признать вторичными. А это означает, что никакого древнейшего безмитохондриального этапа в истории эукариот — по крайней мере, их современных групп — не было. Маргулис считала (для своего времени достаточно обоснованно), что на определенном отрезке истории жизни эукариотизация была широкой тенденцией — «трендом», как сейчас принято говорить. Исходя из этого, вполне можно было бы допустить, что разные эукариоты имеют разных предков: например, что эукариотные водоросли произошли от цианобактерий, животные — от хищных бактерий, а грибы — от бактерий-осмотрофов, всасывающих питательные вещества сквозь поверхность клетки. Никаким фундаментальным законам биологии такая гипотеза не противоречит. Но вот фактам она, к сожалению, противоречит разительно. Молекулярная систематика показывает, что общий предок растений, животных и грибов был не переходной формой, а истинным эукариотом, «полностью оперившимся», как выражается Ник Лейн. Можно смело утверждать, что общий предок всех современных эукариот уже был полноценной эукариотной клеткой: у него было ядро, эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофиламенты, митохондрии и жгутики. В общем, полный набор эукариотных признаков. Обратим внимание, что в этот набор признаков не входят хлоропласты. Они появились далеко не у всех эукариот и не сразу. Кроме того, хлоропласты уж точно приобретались неоднократно, причем разными способами в разных эволюционных ветвях. Хлоропласты бывают как первичные (когда эукариот захватывает цианобактерию), так и вторичные (когда эукариот захватывает другого эукариота с цианобактерией внутри) и даже третичные (когда один эукариот захватывает второго эукариота, внутри которого живет третий эукариот, а уж внутри того — цианобактерия). Здесь эволюция, что называется, разгулялась. С митохондриями ситуация совершенно иная: по признаку их наличия мы не видим никакого особого разнообразия и никаких переходных стадий (если не считать многочисленных фактов вторичной потери, но о происхождении эукариот такие факты не говорят ровно ничего). Если бы сценарий Маргулис был полностью верен, то и с митохондриями, и со жгутиками дело обстояло бы примерно так же, как с хлоропластами, — но этого нет. В чем Маргулис была права, так это в том, что эукариоты в целом весьма предрасположены к захвату эндосимбионтов. Тут можно привести самые разные примеры, вплоть до приобретения некоторыми глубоководными червями симбионтов-бактерий, за счет которых эти черви, собственно, и живут (В. В. Малахов, 1997. Вестиментиферы — автотрофные животные). Бурная эволюция хлоропластов — самое яркое проявление этой тенденции. Только вот «действующие лица», которые их приобрели, по-видимому, уже имели к тому времени полный набор эукариотных признаков, включая митохондрии. Конфигурация эволюционного древа эукариот, насколько мы ее сейчас знаем, просто не допускает других версий. К этому Лейн добавляет, что базовая структура клеток на удивление мало отличается у разных эукариот в зависимости от их образа жизни (хотя сам образ жизни может отличаться очень сильно). Все характерные компоненты клетки, делающие ее эукариотной, устроены в целом одинаково и у растений, и у животных, и у грибов, и у жгутиконосцев, и у амеб... «Мы теперь знаем, что почти все различия между эукариотами отражают вторичные адаптации», — пишет Лейн в обсуждаемой статье. Единообразие устройства эукариотной клетки означает, что первые этапы ее становления не оставили в современном разнообразии эукариот практически никаких следов. Уникальное событиеВыводы, которые делает Лейн, на сегодняшний день уже нельзя назвать новыми или неожиданными. Современные данные наиболее совместимы с предположением, что становление эукариотной клетки было единичным событием, завершившимся (в доступном нам масштабе времени) очень быстро. Вероятно, предки эукариот прошли на этом этапе через своего рода «бутылочное горлышко» (в одной более ранней статье Лейн предполагал, что это была маленькая неустойчивая короткоживущая популяция, в которой и свершились все основные перемены; N. Lane, 2011. Energetics and genetics across the prokaryote-eukaryote divide). В результате возник «полностью оперившийся» первый эукариот, потомки которого разошлись по разным экологическим нишам — но фундаментальное устройство клетки у них уже не менялось. Никакой параллельной эукариотизации, таким образом, не было. Во всяком случае, современная биология не находит подтверждающих ее свидетельств. Данные сравнительной геномики позволяют предположить, что пороговым событием, выделившим эукариот из всей остальной живой природы, было объединение двух клеток — архейной (вероятно, принадлежавшей кому-то из локиархеот) и бактериальной (вероятно, принадлежавшей кому-то из протеобактерий). Образовавшийся суперорганизм и стал первым эукариотом (рис. 3, Б). Современная «мэйнстримная» точка зрения отождествляет это событие с приобретением митохондрий (так называемый «раннемитохондриальный» сценарий; см., например: N. Yutin et al., 2009. The origins of phagocytosis and eukaryogenesis). Действительно, митохондрии — бесспорные потомки протеобактерий, и они-то уж точно проникли в качестве симбионтов в клетку археи (либо примитивного эукариота, не слишком далеко ушедшего от архей). Правда, на вопрос о том, как именно они туда попали, Лейн дает довольно неожиданный ответ. А именно: «Мы не знаем». В чем тут дело? Согласно классической теории, все внутренние симбионты были приобретены эукариотными клетками путем фагоцитоза, то есть захвата ложноножками с изоляцией захваченного объекта и последующим его перевариванием (в данном случае — несостоявшимся). В отношении хлоропластов это, по всей видимости, верно, а вот в отношении митохондрий — очень сомнительно. Предположение, что фагоцитоз появился раньше, чем митохондрии, плохо согласуется с данными биоинформатики. Сравнительный анализ белковых последовательностей показывает, что актиновые микрофиламенты, образующие внутренний каркас любых ложноножек, скорее всего, сначала были неподвижными — белки, позволяющие им еще и сокращаться, появились заметно позже (Е. В. Кунин, 2014. Логика случая). А это означает, что начаться прямо с фагоцитоза эволюция эукариот не могла — митохондрии были приобретены каким-то другим способом. Здесь открывается большой простор для гипотез. Есть, например, весьма изящно обоснованное предположение, что архейная клетка не поглощала предков митохондрий путем фагоцитоза, а охватила их своими постепенно разраставшимися выростами, которые в конце концов сомкнулись, изолировав митохондрии от внешней среды и заодно создав систему внутриклеточных полостей — то, что сейчас называется эндоплазматической сетью (D. Baum, B. Baum, 2014. An inside-out origin for the eukaryotic cell). Есть другая остроумная гипотеза: предки митохондрий были внутриклеточными паразитами (Z. Wang, M. Wu, 2014. Phylogenomic reconstruction indicates mitochondrial ancestor was an energy parasite). Авторы этой последней идеи обратили внимание, что в группе альфа-протеобактерий, из которой, несомненно, вышли митохондрии, есть немало специализированных внутриклеточных паразитов — например, риккетсии. Проведенная биоинформатическими методами реконструкция вероятного предка митохондрий показывает, что это была бактерия с маленьким геномом, неспособная самостоятельно синтезировать некоторые аминокислоты и компоненты нуклеотидов. Эти жизненно важные молекулы она всасывала из внешней среды с помощью специальных транспортных белков. Такой образ жизни в высшей степени характерен для внутриклеточных паразитов, зависимых от хозяйского обмена веществ. Более того, предок митохондрий, скорее всего, имел встроенный во внешнюю мембрану АТФ/АДФ-антипортер (см. ATP/ADP antiporter) — белок, способный выкачивать из внешней среды аденозинтрифосфат (АТФ), вещество, которое используется всеми живыми клетками как универсальный носитель энергии. Многие внутриклеточные паразиты, в том числе и риккетсии, используют этот белок для «воровства» энергии (в форме АТФ) из клетки хозяина. Существа, ведущие такой образ жизни, называются энергетическими паразитами. Авторы упомянутой гипотезы считают, что к ним относился и предок митохондрий — бактерия, которая первоначально паразитировала внутри чужих клеток. Дело в том, что механизм АТФ/АДФ-антипортера можно «раскрутить» в другую сторону, и тогда он начнет отдавать АТФ во внешнюю среду вместо того, чтобы выкачивать его оттуда. В ходе симбиоза это могло бы стать хорошей основой для смены функций, превратившей предков митохондрий из энергетических паразитов в поставщиков энергии — разумеется, не бесплатной. Но надо подчеркнуть, что всё это пока лишь предположения. Загадка происхождения митохондрий, не говоря уж о происхождении ядра, до сих пор не разгадана. Случайность и необходимостьИтак, верна ли гипотеза сериального эндосимбиоза? Да — в том смысле, что в истории эукариот действительно много раз случались симбиотические события. Лучше всего это иллюстрирует долгая, богатая и неплохо сейчас изученная история хлоропластов (P. Keeling et al., 2013. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution). Нет — в том смысле, что сериальный эндосимбиоз не был предпосылкой возникновения эукариот как группы. Эндосимбиотическое событие, которое привело к возникновению эукариот, было, насколько мы сейчас можем судить, уникальным. Таким образом, сценарий «параллельной эукариотизации» не подтверждается. Это отнюдь не значит, что эволюционных событий подобного типа вообще не бывает: некоторые из них подробно описаны палеонтологами (например, маммализация зверообразных рептилий, которые приобретают признаки млекопитающих параллельно в нескольких эволюционных ветвях). Более того, список подобных «параллельных сценариев» в последнее время даже пополняется. «Элементы» не раз писали о гипотезе независимого возникновения нервной системы в двух совершенно разных ветвях многоклеточных животных (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Но возникновение эукариот — одно из самых уникальных событий во всей истории жизни на Земле. Вероятно, потому оно и выпадает из этого ряда. В современной научной литературе есть такое понятие, как гипотеза редкой Земли (см. Rare Earth hypothesis). Сторонники этой гипотезы допускают, что относительно просто устроенная жизнь (бактериального уровня организации) может существовать на множестве планет и быть во Вселенной довольно обычным явлением. А вот относительно сложная жизнь (эукариотная или сопоставимая с ней) возникает только при редчайшем стечении обстоятельств; не исключено, что планета с подобной жизнью — всего одна в Галактике. Если гипотеза редкой Земли верна, то именно возникновение эукариот, скорее всего, является рубежным событием, отделяющим «простую» жизнь (широко распространенную) от «сложной» (маловероятной). К похожим выводам недавно (и совершенно независимо) пришел автор известной книги «Происхождение жизни» Михаил Никитин. «Пока мы не знаем даже, насколько закономерно было появление эукариот. Если для других этапов развития жизни, таких как переход от мира РНК к РНК-белковому миру, обособление прокариотных клеток из доклеточного “мира вирусов” или появление фотосинтеза, мы с уверенностью можем сказать, что они закономерны и практически неизбежны, коль скоро жизнь уже появилась, то появление эукариот в прокариотной биосфере могло быть очень маловероятно. Возможно, что в нашей Галактике есть миллиарды планет с жизнью бактериального уровня, но только на Земле появились эукариоты, на основе которых появились многоклеточные животные и затем разумные существа» (М. Никитин, 2014. Выдвинута новая гипотеза происхождения эукариотической клетки). Может быть, нам потому так и сложно разобраться в деталях происхождения эукариот: это уникальное (в масштабах планеты) событие, к которому очень трудно приложить принцип униформизма, требующий «по умолчанию» исходить из единообразия факторов и процессов во все моменты времени. Но как раз поэтому загадка происхождения эукариот — одна из самых увлекательных во всей биологии. Нерешенных вопросов в этой области еще множество, здесь (как и в обсуждаемой статье Ника Лейна) упомянуты далеко не все из них. Источник: Nick Lane. Serial endosymbiosis or singular event at the origin of eukaryotes? // Journal of Theoretical Biology. 7 December 2017. V. 434. P. 58–67. Сергей Ястребов |
Рис. 1. Пути эволюции древнейших эукариот, согласно взглядам Линн Маргулис. Здесь показаны три симбиогенетических события — приобретение митохондрий, жгутиков и хлоропластов, — которые могли происходить в разных эволюционных ветвях в разном порядке. Тем не менее «верхний» и «нижний» пути эволюции ведут к одному и тому же состоянию: клетке, имеющей и ядро (происхождение которого тут оставлено «за кадром»), и митохондрии, и жгутики. У предков растений к этому добавляются еще и хлоропласты. Прокариотами называются все клеточные организмы, кроме эукариот, архезоями — гипотетические эукариоты, никогда не имевшие митохондрий. Иллюстрация с сайта earthstep.wordpress.com, с изменениями